A embalagem de DNA refere-se ao processo pelo qual o DNA é compactado e organizado dentro do núcleo celular. O DNA é uma molécula longa e fina que contém as informações genéticas de um organismo. Para que o DNA possa caber dentro do núcleo celular, ele precisa ser condensado em estruturas mais compactas, como os cromossomos. A embalagem do DNA envolve a interação com proteínas chamadas histonas, que ajudam a enrolar e compactar o DNA em uma estrutura mais densa. Esse processo de embalagem do DNA é fundamental para regular a expressão gênica e garantir a estabilidade do material genético.
Entenda a função do DNA e sua importância para os seres vivos.
O DNA, ou ácido desoxirribonucleico, é uma molécula presente em todas as células dos seres vivos, responsável por armazenar e transmitir informações genéticas. Ele é essencial para a vida, pois contém as instruções necessárias para o desenvolvimento, funcionamento e reprodução dos organismos.
O DNA é composto por uma sequência de nucleotídeos, que formam os genes. Cada gene é responsável por uma característica específica, como a cor dos olhos, a altura, a forma do corpo, entre outras. A combinação dos genes determina as características de um indivíduo e influencia seu comportamento e saúde.
A função do DNA é codificar proteínas, que são essenciais para a estrutura e funcionamento das células. Sem as proteínas, as células não conseguiriam desempenhar suas funções vitais, o que levaria à morte do organismo. Portanto, o DNA é fundamental para a sobrevivência e a reprodução dos seres vivos.
A embalagem de DNA, ou cromatina, é uma estrutura formada por proteínas que ajudam a compactar e organizar o DNA dentro do núcleo celular. Essa organização é importante para garantir que o DNA seja protegido e acessado de maneira eficiente durante os processos celulares, como a replicação e a transcrição.
Em resumo, o DNA é crucial para a vida dos seres vivos, pois contém as informações genéticas necessárias para a sobrevivência e reprodução. A embalagem de DNA ajuda a manter a integridade e a funcionalidade dessa molécula fundamental para a existência de todos os organismos.
O que é DNA e sua definição em poucas palavras explicadas de forma simplificada.
O DNA é uma molécula presente em todas as células dos seres vivos e que contém as informações genéticas necessárias para o funcionamento e desenvolvimento do organismo. Ele é composto por uma sequência de nucleotídeos que formam os genes responsáveis por características hereditárias. Em resumo, o DNA é como um manual de instruções que determina as características de um organismo.
A embalagem de DNA refere-se ao processo em que o DNA é organizado e compactado dentro do núcleo das células. Isso é necessário para que a molécula de DNA caiba dentro do núcleo e possa se desenrolar e ser acessada quando necessário para a transcrição de genes. A embalagem de DNA envolve a formação de estruturas como cromossomos, que são compostos por DNA e proteínas histonas.
Conhecendo os diferentes tipos de DNA: uma visão geral sobre sua diversidade genética.
A embalagem de DNA refere-se ao processo pelo qual o DNA é compactado e organizado dentro do núcleo das células. O DNA é uma molécula longa e fina que contém toda a informação genética de um organismo. Para que o DNA caiba dentro do núcleo celular, ele precisa ser embalado de forma eficiente.
Existem diferentes tipos de DNA, cada um com sua própria função e estrutura. O DNA genômico, por exemplo, é o tipo de DNA encontrado no núcleo das células e contém todos os genes de um organismo. Já o DNA mitocondrial é encontrado nas mitocôndrias e é responsável pela produção de energia nas células.
Além disso, o DNA também pode ser classificado de acordo com sua sequência de nucleotídeos. Por exemplo, o DNA de um organismo pode ser classificado como DNA codificante, que contém os genes que são expressos para produzir proteínas, ou como DNA não codificante, que regula a expressão dos genes.
Em resumo, conhecer os diferentes tipos de DNA nos permite ter uma visão mais ampla da diversidade genética presente nos organismos vivos. A embalagem de DNA é um processo fundamental para garantir que a informação genética seja armazenada e transmitida de forma eficiente em todas as células de um organismo.
Significado da sigla DNA: Descubra o que representa essa importante abreviação científica.
O DNA, sigla para Ácido Desoxirribonucleico, é uma molécula presente em todos os seres vivos. Essa substância é responsável por armazenar e transmitir as informações genéticas de um organismo, determinando suas características físicas e biológicas.
Quando falamos em embalagem de DNA, nos referimos à forma como essa molécula é organizada dentro das células. O DNA é compactado em estruturas chamadas cromossomos, que garantem que a informação genética seja protegida e possa ser replicada de forma eficiente durante a divisão celular.
A embalagem de DNA é fundamental para garantir a estabilidade e integridade do material genético de um organismo. Sem uma organização adequada, as informações contidas no DNA poderiam se perder ou sofrer mutações, o que poderia levar a problemas genéticos e até mesmo doenças.
Portanto, a embalagem de DNA é um processo essencial para a manutenção da vida e da diversidade biológica em nosso planeta. É graças a essa organização cuidadosa que os seres vivos conseguem se desenvolver e se reproduzir de forma saudável e funcional.
O que é embalagem de DNA?
A embalagem de ADN é um termo que define a compactação controlada do DNA na célula. Em nenhuma célula (e de fato, nem mesmo em vírus), o DNA é livre, relaxado e em verdadeira solução.
O DNA é uma molécula extremamente longa que, além disso, está sempre interagindo com uma enorme variedade de proteínas diferentes. Para o processamento, herança e controle da expressão dos genes que ele carrega, o DNA adota uma organização espacial específica. Isso é alcançado pela célula, controlando rigorosamente cada etapa da embalagem do DNA em diferentes níveis de compactação.
Os vírus têm estratégias de empacotamento diferentes para seus ácidos nucleicos. Um dos favoritos é a formação de espirais compactas. Pode-se dizer que os vírus são ácidos nucleicos empacotados nas proteínas que os cobrem, protegem e mobilizam.
Nos procariontes, o DNA está associado a proteínas que determinam a formação de ligações complexas em uma estrutura chamada nucleoide. O nível máximo de compactação de DNA em uma célula eucariótica, por outro lado, é o cromossomo mitótico ou meiótico.
O único exemplo em que um DNA-B não é empacotado é um laboratório de pesquisa que persegue esse objetivo.
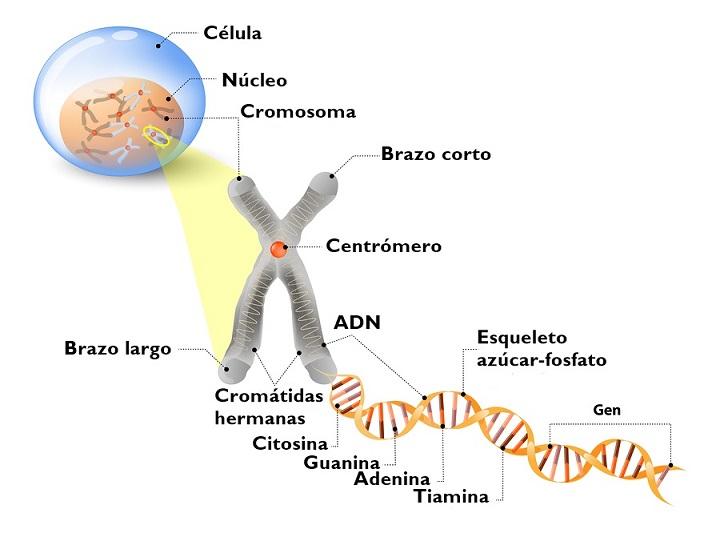
Estrutura de DNA

O DNA é formado por duas bandas antiparalelas que formam uma dupla hélice. Cada um deles possui um esqueleto de ligações fosfodiéster nas quais açúcares ligados a bases de nitrogênio estão ligados.
Dentro da molécula, as bases de nitrogênio de uma banda formam ligações de hidrogênio (duas ou três) com a banda complementar.
Em uma molécula como essa, os ângulos de ligação mais importantes mostram rotação livre. As ligações de açúcar nitrogenado, grupo açúcar-fosfato e ligação fosfodiéster são flexíveis.
Isso permite que o DNA, visto como uma haste flexível, mostre alguma capacidade de dobrar e enrolar. Essa flexibilidade permite ao DNA adotar estruturas locais complexas e formar laços de interação a curta, média e longa distância.
Essa flexibilidade também explica como 2 metros de DNA podem ser mantidos em cada célula diplóide de um ser humano. Em um gameta (célula haplóide), seria um metro de DNA.
Nucleoide bacteriano
Embora não seja uma regra inquebrável, o cromossomo bacteriano existe como uma única molécula de DNA de banda dupla superenrolada.
A dupla hélice gira mais sobre si mesma (mais de 10 pb por turno), produzindo alguma compactação. Nós locais também são gerados graças a manipulações que são enzimaticamente controladas.
Além disso, existem seqüências no DNA que permitem a formação de domínios em grandes ligações. Chamamos de estrutura resultante do super-resfriamento e de laços ordenados nucleoide.
Estes sofrem mudanças dinâmicas graças a algumas proteínas que fornecem alguma estabilidade estrutural ao cromossomo compactado. O grau de compactação em bactérias e arquéias é tão eficiente que pode haver mais de um cromossomo por nucleoide.
O nucleoide compacta no DNA procariótico pelo menos cerca de 1000 vezes. A estrutura topológica do nucleoide é uma parte fundamental da regulação dos genes que o cromossomo transporta. Ou seja, estrutura e função constituem a mesma unidade.
Níveis de compactação de cromossomos eucarióticos
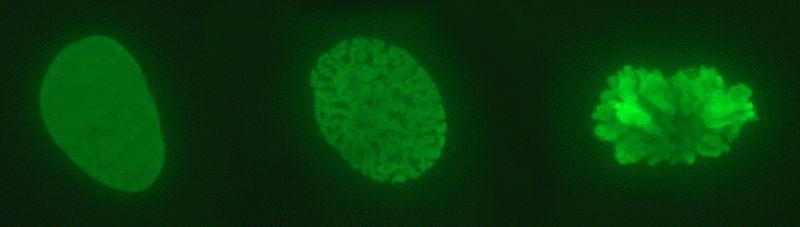
O DNA no núcleo eucariótico não está nu. Interage com muitas proteínas, das quais as mais importantes são histonas. As histonas são pequenas proteínas carregadas positivamente que se ligam ao DNA de maneira não específica.
O que observamos no núcleo é um complexo de DNA: histonas, que chamamos de cromatina. Cromatina altamente condensada, que geralmente não é expressa, é heterocromatina. Pelo contrário, a menos compactada (mais relaxada), ou eucromatina, é a cromatina com genes que são expressos.
A cromatina tem vários níveis de compactação. O mais elementar é o do nucleossomo; Eles são seguidos pelas ligações da fibra solenóide e da cromatina em interfase. Somente quando um cromossomo se divide é que os níveis máximos de compactação são mostrados.
Nucleossomo
O nucleossomo é a unidade básica da organização da cromatina. Cada nucleossomo é formado por um octâmero de histonas que formam uma espécie de tambor.
O octâmero consiste em duas cópias de cada uma das histonas H2A, H2B, H3 e H4. Ao redor deles, o DNA leva quase 1,7 turnos. É seguido por uma fração de DNA livre denominada ligante de 20 pb associado à histona H1 e depois outro nucleossomo. A quantidade de DNA em um nucleossomo e a que o liga a outro é de cerca de 166 pares de bases.
Esta etapa de empacotamento do DNA compacto para a molécula cerca de 7 vezes. Ou seja, passamos de um metro para pouco mais de 14 cm de DNA.
Esse empacotamento é possível porque as histonas positivas cancelam a carga negativa do DNA e a consequente autorepulsão eletrostática. A outra razão é que o DNA pode ser dobrado de maneira a girar o octâmero da histona.
Fibra de 30 nm
A fibra de contas em um colar que forma muitos nucleossomos sucessivos é enrolada em uma estrutura mais compactada.
Embora não tenhamos certeza de qual estrutura ela realmente adota, sabemos que ela atinge uma espessura de cerca de 30 nm. Essa é a chamada fibra de 30 nm; A histona H1 é essencial para a sua formação e estabilidade.
A fibra de 30 nm é a unidade estrutural básica da heterocromatina. O dos nucleossomos frouxos, o da eucromatina.
Laços e voltas
A fibra de 30 nm, no entanto, não é completamente linear. Pelo contrário, forma alças de cerca de 300 nm de comprimento, de maneira sinuosa, em uma matriz protéica pouco conhecida.
Essas ligações em uma matriz proteica formam uma fibra cromatina mais compacta de 250 nm de diâmetro. Finalmente, eles estão alinhados como uma simples hélice de 700 nm de espessura, dando origem a uma das cromátides irmãs de um cromossomo mitótico.
No final, o DNA na cromatina nuclear é compactado cerca de 10.000 vezes no cromossomo da célula em divisão. No núcleo interfásico, sua compactação também é alta, pois é cerca de 1000 vezes comparada ao DNA “linear”.
Compactação de DNA meiótico

No mundo da biologia do desenvolvimento, diz-se que a gametogênese redefine o epigenoma. Ou seja, apaga as marcas de DNA que a vida de quem deu origem ao gameta produziu ou experimentou.
Essas marcas incluem a metilação do DNA e modificações covalentes das histonas (Código da histona). Mas nem todo o epigenoma é redefinido. O que resta das marcas será responsável pela impressão genética paterna ou materna.
A reposição implícita da gametogênese é mais fácil de ver no esperma. No esperma, o DNA não está repleto de histonas. Portanto, as informações associadas às suas modificações no organismo produtor, geralmente, não são herdadas.
Nos espermatozóides, o DNA é empacotado graças à interação com proteínas inespecíficas de ligação ao DNA chamadas protaminas. Essas proteínas formam pontes dissulfeto umas com as outras, contribuindo assim para a sobreposição de camadas de DNA que não se repelem eletrostaticamente.
Referências
- É importante ressaltar que, em alguns casos, é necessário que o paciente tenha um diagnóstico precoce da doença. WW Norton & Company, Nova Iorque, NY, EUA.
- Annunziato, A. (2008) Embalagem de DNA: Nucleossomos e cromatina. Educação da Natureza 1:26. (https://www.nature.com/scitable/topicpage/dna-packaging-nucleosomes-and-chromatin-310).
- Brooker, RJ (2017). Genética: Análise e Princípios. Ensino Superior McGraw-Hill, Nova York, NY, EUA.
- Martínez-Antonio, A. Medina-Rivera, A., Collado-Vides, J. (2009) Mapa estrutural e funcional de um nucleoide bacteriano. Genome Biology, doi: 10.1186 / gb-2009-10-12-247.
- Mathew-Fenn, R. S, Das, R., Harbury, PAB (2008) Remeasuring a dupla hélice. Science, 17: 446-449.
- Travers, AA (2004) A base estrutural da flexibilidade do DNA. Transações Filosóficas da Sociedade Real de Londres, Série A, 362: 1423-1438.
- Travers, A., Muskhelishvili, G. (2015) Estrutura e função do DNA. FEBS Journal, 282: 2279-2295.