O Ciclo Cori é um processo metabólico que ocorre no fígado durante a gliconeogênese para produzir glicose a partir de lactato e alanina. Esse ciclo desempenha um papel importante na regulação dos níveis de glicose no sangue durante o jejum ou exercício físico intenso. Neste ciclo, o lactato e a alanina são convertidos em glicose através de uma série de reações enzimáticas. Este processo é vital para fornecer energia ao organismo em momentos de demanda elevada, garantindo a manutenção dos níveis de glicose necessários para o funcionamento adequado do corpo. Neste artigo, abordaremos as etapas e recursos envolvidos no Ciclo Cori, fornecendo uma visão geral deste importante processo metabólico.
Entendendo o processo de regeneração de glicose a partir do ácido lático.
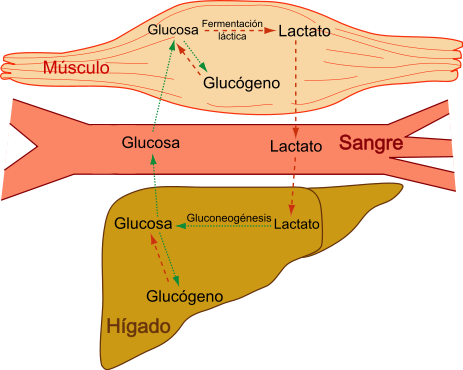
O processo de regeneração de glicose a partir do ácido lático é conhecido como Ciclo Cori. Esse ciclo ocorre no fígado e é essencial para a manutenção dos níveis de glicose no organismo. Quando há uma demanda por energia e a glicose não está disponível em quantidade suficiente, o ácido lático produzido durante a glicólise nos músculos é transportado para o fígado.
No fígado, o ácido lático é convertido de volta em glicose por meio de uma série de reações bioquímicas. O ácido lático é primeiro convertido em piruvato e, em seguida, o piruvato é convertido em glicose por um processo conhecido como neoglicogênese. Essa glicose é então liberada na corrente sanguínea para ser utilizada pelos tecidos que necessitam de energia.
O Ciclo Cori é essencial em situações de demanda energética, como durante exercícios físicos intensos ou em casos de hipoglicemia. Ele permite que o organismo regenere glicose a partir do ácido lático, garantindo um suprimento constante de energia para as células.
É importante ressaltar que o Ciclo Cori é um processo energicamente custoso, pois a regeneração de glicose a partir do ácido lático requer gasto de energia. No entanto, é fundamental para a manutenção do equilíbrio energético do organismo e para garantir o funcionamento adequado dos tecidos em momentos de necessidade.
Qual é a função metabólica do ciclo de Cori no organismo humano?
O ciclo de Cori é um processo metabólico fundamental para o organismo humano, principalmente durante atividades físicas intensas ou em situações de baixo consumo de oxigênio. Ele desempenha um papel crucial na regulação dos níveis de glicose no sangue e na produção de energia para as células.
Este ciclo ocorre principalmente no fígado e nos músculos esqueléticos. Durante a contração muscular, ocorre a quebra da glicose para produzir energia na forma de ATP. Como resultado, há um acúmulo de ácido lático nos músculos, o que pode causar fadiga e desconforto.
O ciclo de Cori entra em ação para converter o ácido lático produzido nos músculos de volta em glicose no fígado. Essa glicose é então liberada na corrente sanguínea para ser utilizada como combustível pelas células, incluindo os músculos que estão em atividade.
Assim, a principal função metabólica do ciclo de Cori é reciclar o ácido lático em glicose para garantir um suprimento contínuo de energia durante exercícios físicos intensos. Além disso, ele contribui para manter os níveis adequados de glicose no sangue, prevenindo hipoglicemia e garantindo o funcionamento adequado do organismo.
Em resumo, o ciclo de Cori é essencial para a regulação do metabolismo de glicose e para a produção de energia durante atividades físicas intensas, garantindo o bom funcionamento do organismo humano.
Entenda os processos metabólicos dos ciclos de Cori e alanina na produção de energia.
O ciclo de Cori e o ciclo da alanina são dois processos metabólicos essenciais para a produção de energia no organismo. Esses ciclos permitem a regeneração do NAD+ e a produção de glicose a partir do lactato, contribuindo para a manutenção dos níveis de glicose no sangue durante períodos de alta demanda energética.
No ciclo de Cori, o lactato produzido durante a glicólise nos músculos esqueléticos é transportado para o fígado, onde é convertido de volta em glicose através da gliconeogênese. Esse processo requer a utilização de energia na forma de ATP, mas permite a regeneração do NAD+ necessário para a continuidade da glicólise nos músculos.
Já no ciclo da alanina, o grupo amino da alanina é transferido para o alfa-cetoglutarato, formando piruvato e glutamato. O piruvato pode então ser convertido em glicose no fígado através da gliconeogênese, enquanto o glutamato é convertido em alanina nos músculos para ser transportado de volta ao fígado.
Esses ciclos são fundamentais para garantir um suprimento constante de glicose para os tecidos que dependem desse substrato para a produção de energia, como o cérebro e os glóbulos vermelhos. Além disso, eles contribuem para a manutenção do equilíbrio ácido-base no organismo, ao eliminar o excesso de lactato produzido durante exercícios intensos.
Portanto, compreender os processos metabólicos dos ciclos de Cori e alanina é essencial para entender como o organismo regula a produção de energia e mantém a homeostase durante diferentes condições fisiológicas.
Entenda o processo de formação e utilização do lactato durante a atividade física.
O lactato é um subproduto do metabolismo anaeróbio que ocorre durante a atividade física intensa. Quando os músculos não recebem oxigênio suficiente para produzir energia, eles começam a quebrar a glicose para obter ATP, resultando na formação de lactato.
Este lactato é então liberado na corrente sanguínea e transportado para o fígado, onde é convertido de volta em glicose através de um processo chamado Ciclo Cori. Essa glicose pode ser utilizada novamente pelos músculos como fonte de energia durante a atividade física.
O Ciclo Cori consiste em várias etapas, incluindo a conversão de lactato em piruvato no fígado, a conversão de piruvato em glicose e a liberação de glicose na corrente sanguínea para ser utilizada pelos músculos. Este ciclo é essencial para manter os níveis de glicose e energia durante a atividade física intensa.
Portanto, o lactato não é apenas um subproduto prejudicial do metabolismo anaeróbio, mas também desempenha um papel importante na regulação dos níveis de energia durante a atividade física. Entender o processo de formação e utilização do lactato, juntamente com o Ciclo Cori, é fundamental para otimizar o desempenho físico e a recuperação após o exercício.
Ciclo Cori: etapas e recursos
O ciclo Cori ou ciclo do ácido láctico é uma via metabólica na qual o lactato produzido pelas vias glicolíticas no músculo vai para o fígado, onde é convertido novamente em glicose. Este composto retorna ao fígado para ser metabolizado.
Essa via metabólica foi descoberta em 1940 por Carl Ferdinand Cori e sua esposa Gerty Cori, cientistas da República Tcheca. Ambos ganharam o Prêmio Nobel de fisiologia ou medicina.
Processo (etapas)
Glicólise muscular anaeróbica
O ciclo Cori começa nas fibras musculares. Neste tecido, a obtenção de ATP ocorre principalmente pela conversão de glicose em lactato.
Deve-se mencionar que os termos ácido lático e lactato, amplamente utilizados na terminologia esportiva, diferem ligeiramente em sua estrutura química. O lactato é o metabolito produzido pelos músculos e é a forma ionizada, enquanto o ácido lático possui um próton adicional.
A contração muscular ocorre por hidrólise de ATP.
Isso é regenerado por um processo chamado “fosforilação oxidativa”. Essa rota ocorre nas mitocôndrias das fibras musculares de contração lenta (vermelha) e rápida (branca)
As fibras musculares rápidas são compostas de miosinas rápidas (40-90 ms), em contraste com as fibras do cristalino, formadas por miosinas lentas (90-140 ms). Os primeiros produzem mais força, mas rapidamente fadiga.
Gliconeogênese no fígado
Através do sangue, o lactato atinge o fígado. Novamente, o lactato é convertido em piruvato pela ação da enzima lactato desidrogenase.
Finalmente, o piruvato é transformado em glicose por gliconeogênese, utilizando ATP hepático, gerado por fosforilação oxidativa.
Essa nova glicose pode retornar ao músculo, onde é armazenada na forma de glicogênio e usada novamente para contração muscular.
Reações de gliconeogênese
Gliconeogênese é a síntese de glicose usando componentes não- carboidratos . Esse processo pode levar piruvato, lactato, glicerol e a maioria dos aminoácidos como matéria-prima.
O processo começa nas mitocôndrias, mas a maioria das etapas continua no citosol celular.
A gliconeogênese envolve dez das reações da glicólise, mas na direção oposta. Ocorre da seguinte maneira:
-Na matriz mitocondrial, o piruvato é convertido em oxaloacetato por meio da enzima piruvato carboxilase. Esta etapa precisa de uma molécula de ATP, que se torna ADP, uma molécula de CO 2 e uma molécula de água. Essa reação libera dois H + para o meio.
-O oxaloacetato é convertido em l-malato pela enzima malato desidrogenase. Essa reação precisa de uma molécula de NADH e H.
-L-malato deixa o citosol onde o processo continua. O malato volta ao oxaloacetato. Esta etapa é catalisada pela enzima malato desidrogenase e envolve o uso de uma molécula de NAD +
-O oxaloacetato é convertido em fosfoenolpiruvato pela enzima fosfoenolpiruvato carboxiquinase. Esse processo envolve uma molécula de GTP que passa para o PIB e o CO 2 .
-O fosfoenolpiruvato passa para o 2-fosfoglicerato pela ação da enolase. Esta etapa requer uma molécula de água.
– A fosfoglicerato mutase catalisa a conversão de 2-fosfoglicerato em 3-fosfoglicerato.
O -3-fosfoglicerato passa para o 1,3-bisfosfoglicerato, catalisado pela fosfoglicerato mutase. Esta etapa requer uma molécula de ATP.
-O 1,3-bifosfoglicerato é catalisado em d-gliceraldeído-3-fosfato pela gliceraldeído-3-fosfato desidrogenase. Esta etapa envolve uma molécula de NADH.
-D-gliceraldeído-3-fosfato passa para 1,6-bifosfato de frutose através da aldolase.
– O 1,6-bifosfato de frutose é convertido em 6-fosfato de frutose pela 1,6-bifosfatase de frutose. Essa reação envolve uma molécula de água.
– O 6-fosfato de frutose é convertido em 6-fosfato de glicose pela enzima glicose-6-fosfato isomerase.
-Finalmente, a enzima glicose 6-fosfatase catalisa a passagem deste último composto para α-d-glicose.
Por que o lactato precisa viajar para o fígado?
As fibras musculares não são capazes de realizar o processo de gliconeogênese. Se eu pudesse, seria um ciclo totalmente injustificado, pois a gliconeogênese usa muito mais ATP do que a glicólise.
Além disso, o fígado é um tecido apropriado para o processo. Nesse órgão, ele sempre tem a energia necessária para realizar o ciclo, porque não há falta de O 2 .
Tradicionalmente, pensava-se que durante a recuperação celular após o exercício, cerca de 85% do lactato era removido e enviado ao fígado. Então ocorre a conversão em glicose ou glicogênio.
No entanto, novos estudos utilizando ratos como organismo modelo revelam que o destino frequente do lactato é a oxidação.
Além disso, diferentes autores sugerem que o papel do ciclo Cori não é tão significativo quanto se acreditava. Segundo essas investigações, o papel do ciclo é reduzido para apenas 10 ou 20%.
Ciclo de Cori e exercício
Ao se exercitar, o sangue atinge um acúmulo máximo de ácido lático, após cinco minutos de treinamento. Este tempo é suficiente para o ácido lático migrar dos tecidos musculares para o sangue.
Após a fase de treinamento muscular, os níveis de lactato no sangue retornam ao normal após uma hora.
Ao contrário da crença popular, o acúmulo de lactato (ou o próprio lactato) não é a causa da depleção muscular. Foi demonstrado que em exercícios onde o acúmulo de lactato é baixo, ocorre fadiga muscular.
Pensa-se que a verdadeira causa é a diminuição do pH nos músculos. É possível que o pH diminua da linha de base de 7,0 para 6,4, considerado um valor bastante baixo. De fato, se o pH permanecer próximo de 7,0, mesmo que a concentração de lactato seja alta, o músculo não se cansa.
No entanto, o processo que leva à fadiga como resultado da acidificação ainda não está claro. Pode estar relacionado à precipitação de íons cálcio ou a uma diminuição na concentração de íons potássio.
Os atletas recebem massagens e aplicam gelo nos músculos para promover a passagem de lactato para o sangue.
O ciclo da alanina
Existe um caminho metabólico quase idêntico ao ciclo Cori, chamado ciclo alanina. Aqui o aminoácido é o precursor da gliconeogênese. Em outras palavras, a alanina substitui a glicose.
Referências
- Baechle, TR e Earle, RW (Eds.). (2007). Princípios de treinamento de força e fitness . Pan-American Medical Ed.
- Campbell, MK, & Farrell, SO (2011). Bioquímica . Sexta edição . Thomson Brooks / Cole.
- Koolman, J. & Röhm, KH (2005). Bioquímica: texto e atlas . Pan-American Medical Ed.
- Mougios, V. (2006). Exercer bioquímica . Cinética Humana
- Poortmans, JR (2004). Princípios de bioquímica do exercício . 3 rd , edição revisada . Karger
- Voet, D. & Voet, JG (2006). Bioquímica . Pan-American Medical Ed.