A lei do dízimo ecológico, também conhecida como a lei dos 10%, propõe que 10% da renda de uma pessoa seja destinada a causas ambientais e sustentáveis. Assim como o dízimo religioso, que é uma prática de doação de uma parte do patrimônio para a igreja, o dízimo ecológico busca promover a consciência ecológica e incentivar ações em prol do meio ambiente. Essa prática tem como objetivo estimular a responsabilidade ambiental e contribuir para a preservação e sustentabilidade do planeta.
O mandamento bíblico de contribuir com uma décima parte dos ganhos ao Senhor.
O dízimo é uma prática antiga que tem suas raízes na Bíblia. De acordo com o mandamento bíblico, os fiéis devem contribuir com uma décima parte dos seus ganhos ao Senhor, como forma de reconhecimento e gratidão por tudo o que Ele tem feito por eles. Esta prática é conhecida como a lei do dízimo e é uma forma de demonstrar fidelidade a Deus e sustentar a obra da igreja.
O dízimo não se trata apenas de uma obrigação, mas de um ato de fé e confiança no Senhor. Ao contribuir com uma décima parte dos seus ganhos, os fiéis estão demonstrando que confiam em Deus para suprir todas as suas necessidades e abençoar suas vidas. Além disso, o dízimo também é uma forma de retribuir as bênçãos recebidas e ajudar aqueles que mais precisam.
A lei do dízimo não se restringe apenas a contribuições em dinheiro. O dízimo ecológico, por exemplo, é uma prática que incentiva os fiéis a contribuir com 10% dos seus recursos naturais para a preservação do meio ambiente e a sustentabilidade do planeta. Esta forma de dízimo reflete a responsabilidade dos cristãos em cuidar da criação de Deus e garantir um futuro melhor para as próximas gerações.
Em resumo, o mandamento bíblico de contribuir com uma décima parte dos ganhos ao Senhor é uma prática que visa fortalecer a fé, demonstrar gratidão e ajudar aqueles que precisam. Seja através do dízimo em dinheiro ou do dízimo ecológico, o importante é contribuir com generosidade e amor, confiando que Deus irá abençoar e multiplicar todas as ofertas feitas em Seu nome.
Devo continuar dando o dízimo mesmo estando endividado?
Muitas pessoas se questionam se devem continuar dando o dízimo mesmo estando endividado. Afinal, a lei do dízimo é uma prática religiosa que envolve doar 10% de seus ganhos para a igreja. Mas e se você estiver passando por dificuldades financeiras e suas dívidas estiverem se acumulando?
Primeiramente, é importante ressaltar que a lei do dízimo não é uma obrigação imposta por lei, mas sim uma recomendação feita pela maioria das igrejas. Portanto, não há uma resposta única para essa questão, pois cada pessoa deve avaliar sua situação financeira individualmente.
Se você está enfrentando dificuldades financeiras e suas dívidas estão se acumulando, é essencial priorizar o pagamento dessas dívidas para evitar maiores problemas no futuro. Nesse caso, pode ser prudente suspender temporariamente a prática de dar o dízimo até que sua situação financeira esteja mais estável.
No entanto, se você acredita firmemente na importância do dízimo e deseja continuar praticando essa lei mesmo estando endividado, é recomendável que você busque orientação financeira para encontrar um equilíbrio entre suas dívidas e suas doações.
Em suma, a decisão de continuar dando o dízimo mesmo estando endividado é pessoal e deve levar em consideração sua situação financeira atual. O mais importante é manter a transparência e a honestidade em suas finanças, buscando sempre o equilíbrio entre suas dívidas e suas doações.
Entendendo o dízimo: por que é importante e como deve ser praticado.
Entendendo o dízimo: muitas pessoas têm dúvidas sobre o que é o dízimo e por que é importante praticá-lo. O dízimo é uma prática antiga que consiste em doar 10% de seus ganhos para a igreja ou para alguma obra de caridade. É uma forma de agradecer a Deus pelas bênçãos recebidas e de contribuir para o sustento da comunidade religiosa.
Praticar o dízimo é importante porque ajuda a manter as atividades da igreja, como a realização de cultos, eventos sociais e obras de caridade. Além disso, o dízimo é uma forma de demonstrar nossa gratidão a Deus e de contribuir para o bem-estar da comunidade.
Como deve ser praticado o dízimo: a prática do dízimo deve ser feita de forma consciente e regular. O ideal é separar 10% de seus ganhos mensalmente e destinar essa quantia para a igreja ou para alguma obra de caridade. É importante fazer essa doação de forma voluntária e com amor no coração, sem esperar nada em troca.
É importante lembrar que o dízimo não é uma obrigação, mas sim uma forma de expressar nossa gratidão a Deus e de contribuir para o bem-estar da comunidade. Quando praticamos o dízimo com fé e generosidade, estamos semeando boas ações que serão recompensadas no futuro.
Em resumo, o dízimo é uma prática importante para os cristãos, pois nos ensina a partilhar nossas bênçãos com os outros e a contribuir para o bem-estar da comunidade. Ao praticarmos o dízimo de forma consciente e regular, estamos demonstrando nossa gratidão a Deus e ajudando a manter as atividades da igreja e as obras de caridade. Portanto, vamos praticar o dízimo com fé e generosidade, sabendo que estamos fazendo a vontade de Deus e contribuindo para um mundo melhor.
Dízimo: obrigação religiosa estabelecida pela lei ou mandamento divino.
O dízimo é uma prática antiga que remonta aos tempos bíblicos, onde os fiéis eram instruídos a dar uma parte de seus rendimentos para a igreja ou para ajudar os mais necessitados. Essa obrigação religiosa é estabelecida pela lei ou mandamento divino, sendo considerada uma forma de contribuição para a manutenção das atividades religiosas e para ajudar a comunidade.
No entanto, nos dias de hoje, surge uma nova interpretação do dízimo, conhecida como dízimo ecológico ou 10%. Neste modelo, os fiéis são encorajados a destinar 10% de seus rendimentos para a preservação do meio ambiente e para ações de sustentabilidade. Essa prática visa conscientizar as pessoas sobre a importância de cuidar do planeta e de adotar hábitos mais sustentáveis em suas vidas.
Assim, o dízimo ecológico se torna uma forma de contribuição não apenas para a igreja, mas também para a sociedade e para o meio ambiente. É uma maneira de demonstrar responsabilidade e comprometimento com a preservação do planeta, cumprindo assim um mandamento divino de cuidar da criação.
Qual é a lei do dízimo ecológico ou 10%?
A lei do dízimo ecológica , legislação ecológica ou 10% levanta como a energia viaja em sua derivação por diferentes níveis tróficos.Também é frequentemente afirmado que esta lei é simplesmente uma conseqüência direta da segunda lei da termodinâmica.
A energia ecológica é uma parte da ecologia que se preocupa em quantificar os relacionamentos que descrevemos acima. Considera-se que Raymond Lindemann (especificamente em seu trabalho seminal de 1942) foi quem estabeleceu os fundamentos dessa área de estudo.
Seu trabalho se concentrou nos conceitos de cadeia e rede trófica e na quantificação da eficiência na transferência de energia entre diferentes níveis tróficos.
Lindemann faz parte da radiação solar incidente ou energia que uma comunidade recebe, capturando as plantas por fotossíntese e continuando a monitorar essa captura e seu uso subsequente por herbívoros (consumidores primários) e depois por carnívoros (consumidores secundários) ) e, finalmente, pelos decompositores.
Qual é a lei do dízimo ecológico?
Após o trabalho pioneiro de Lindemann, assumiu-se que a eficiência de transferência trófica era de cerca de 10%; de fato, alguns ecologistas se referiram a uma lei de 10%. No entanto, desde então, houve várias confusões em relação a esse problema.
Certamente, não existe lei da natureza que resulte precisamente em um décimo da energia entrando em um nível trófico, que é transferido para o próximo.
Por exemplo, uma compilação de estudos tróficos (em ambientes marinhos e de água doce) revelou que a eficiência da transferência por nível trófico variou entre aproximadamente 2 e 24%, embora a média fosse de 10,13%.
Como regra geral, aplicável a sistemas aquáticos e terrestres, pode-se dizer que a produtividade secundária por herbívoros geralmente está localizada aproximadamente, uma ordem de magnitude abaixo da produtividade primária em que está assentada.
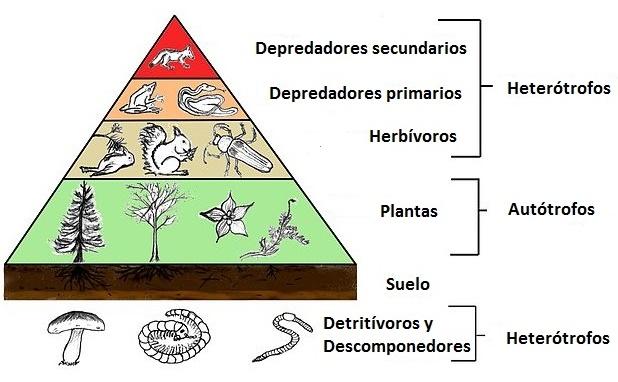
Geralmente, esse relacionamento é mantido em todos os sistemas forrageiros e geralmente se transforma em estruturas piramidais, nas quais a base é fornecida pelas plantas e, nessa base, fica menor, dos consumidores primários, em que outro (menor) consumidor secundário está sentado.
Níveis de organização
Todos os seres vivos requerem matéria e energia; importa para a construção de seus corpos e energia para desempenhar suas funções vitais . Esse requisito não se limita a um organismo individual, mas é extensivo a níveis mais altos de organização biológica que esses indivíduos podem estar em conformidade.
Esses níveis de organização são:
- Uma população biológica : organismos da mesma espécie que vivem na mesma área específica.
- Uma comunidade biológica : conjunto de organismos de diferentes espécies ou populações, que vivem em uma determinada área e interagem através de relações alimentares ou tróficas).
- Um ecossistema : o nível mais complexo de organização biológica, constituído por uma comunidade relacionada ao seu ambiente abiótico – água, luz solar, clima e outros fatores – com os quais interage.
Níveis tróficos
Em um ecossistema, a comunidade e o meio ambiente estabelecem fluxos de energia e matéria.
Os organismos de um ecossistema são agrupados de acordo com um “papel” ou “função” que eles cumprem dentro de cadeias alimentares ou alimentares; É assim que são discutidos os níveis tróficos de produtores, consumidores e decompositores.
Por sua vez, todos e cada um desses níveis tróficos interagem com o ambiente físico-químico que fornece as condições para a vida e, ao mesmo tempo, atua como fonte e fonte de energia e matéria.
Conceitos fundamentais
Produtividade primária grossa e líquida
Primeiro, devemos definir a produtividade primária, que é a taxa na qual a biomassa é produzida por unidade de área.
É geralmente expresso em unidades de energia (Joules por metro quadrado e por dia), ou em unidades de matéria orgânica seca (quilogramas por hectare e por ano) ou como carbono (massa de carbono em kg por metro quadrado por ano).
Em geral, quando nos referimos a toda a energia estabelecida pela fotossíntese, geralmente a chamamos de produtividade primária grossa (PPG).
Disto, uma proporção é gasta na respiração dos mesmos autótrofos (AR) e é perdida como calor. A produção primária líquida (PPN) é obtida subtraindo esse valor do PPG (PPN = PPG-RA).
Essa produção primária líquida (PPN) é a que está finalmente disponível para consumo pelos heterotróficos (são bactérias, fungos e outros animais que conhecemos).
Produtividade secundária
A produtividade secundária (PS) é definida como a taxa de produção de nova biomassa por organismos heterotróficos. Ao contrário das plantas, bactérias, fungos e animais heterotróficos não conseguem produzir compostos complexos e ricos em energia que precisam a partir de moléculas simples.
Eles sempre obtêm sua matéria e energia das plantas, o que podem fazer diretamente consumindo material vegetal ou indiretamente, alimentando-se de outros heterotróficos.
É dessa maneira que plantas ou organismos fotossintéticos em geral (também chamados produtores) compreendem o primeiro nível trófico de uma comunidade; consumidores primários (aqueles que se alimentam de produtores) compõem o segundo nível trófico e consumidores secundários (também chamados carnívoros) compõem o terceiro nível.
Eficiência de transferência e rotas de energia
As proporções da produção primária líquida que fluem ao longo de cada uma das possíveis vias de energia dependem, em última análise, da eficiência da transferência, ou seja, da maneira como a energia é usada e passa de um nível para o outro. outro.
Categorias de eficiência de transferência de energia
Existem três categorias de eficiência na transferência de energia e, com estas bem definidas, podemos prever o padrão do fluxo de energia nos níveis tróficos. Essas categorias são: eficiência de consumo (CE), eficiência de assimilação (EA) e eficiência de produção (EP).
Vamos agora definir essas três categorias mencionadas.
Matematicamente, podemos definir a eficiência de consumo (CE) da seguinte maneira:
CE = I n / P n-1 × 100
Onde podemos ver que CD é uma porcentagem da produtividade total disponível ( P n-1 ) que é efetivamente ingerida pelo compartimento trófico adjacente superior ( I n ).
Por exemplo, para consumidores primários no sistema de pastoreio, CE é a porcentagem (expressa em unidades de energia e por unidade de tempo) do PPN que é consumido pelos herbívoros.
Se estivéssemos nos referindo a consumidores secundários, seria equivalente ao percentual de produtividade de herbívoros consumidos por carnívoros. O resto morre sem ser comido e entra na cadeia de decomposição.
Por outro lado, a eficiência da assimilação é expressa da seguinte forma:
EA = A n / I n × 100
Novamente, nos referimos a uma porcentagem, mas desta vez à parte da energia proveniente dos alimentos e ingerida em um compartimento trófico por um consumidor ( I n ) e que é assimilada por seu sistema digestivo ( A n ).
Essa energia estará disponível para crescimento e para a execução do trabalho. O restante (a parte não assimilada) é perdido com fezes e depois entra no nível trófico dos decompositores.
Finalmente, a eficiência da produção (EP) é expressa como:
EP = P n / A n × 100
que também é uma porcentagem, mas, neste caso, nos referimos à energia assimilada ( A n ) que acaba sendo incorporada à nova biomassa ( P n ). Todo o restante de energia não assimilada é perdido na forma de calor durante a respiração.
Produtos tais como secreções e / ou excreções (energia – rica), que participaram nos processos metabólicos podem ser considerados como produção, P n , e estão disponíveis, como corpos, para decompositores.
Eficiência de transferência global
Tendo definido essas três categorias importantes, agora podemos nos perguntar sobre a “eficiência da transferência global” de um nível trófico para o outro, o que é simplesmente dado pelo produto das eficiências mencionadas acima ( EC x EA x EP ).
Expressos coloquialmente, podemos dizer que a eficiência de um nível é dada pelo que pode ser ingerido efetivamente, que é assimilado e acaba sendo incorporado à nova biomassa.
Para onde vai a energia perdida?
A produtividade dos herbívoros é sempre menor do que a das plantas em que se alimentam. Podemos nos perguntar: para onde vai a energia perdida?
Para responder a essa pergunta, devemos chamar atenção para os seguintes fatos:
- Nem toda biomassa vegetal é consumida pelos herbívoros, pois grande parte morre e entra no nível trófico dos decompositores (bactérias, fungos e outros detritívoros).
- Nem toda a biomassa consumida pelos herbívoros, nem a dos herbívoros consumidos por carnívoros, é assimilada e disponível para ser incorporada na biomassa do consumidor; uma parte é perdida com as fezes e, assim, passa para os decompositores.
- Nem toda energia que é assimilada é realmente convertida em biomassa, pois uma parte é perdida na forma de calor durante a respiração.
Isso acontece por dois motivos básicos: primeiro, porque não há processo de conversão de energia que seja 100% eficiente. Ou seja, sempre há uma perda na forma de calor na conversão, que está em perfeita harmonia com a Segunda Lei da Termodinâmica.
Em segundo lugar, como os animais precisam de trabalho, o que requer gasto de energia e, por sua vez, implica novas perdas na forma de calor.
Esses padrões ocorrem em todos os níveis tróficos e, como previsto pela Segunda Lei da Termodinâmica, parte da energia que se tenta transferir de um nível para outro, sempre se dissipa na forma de calor inutilizável.
Referências
- Caswell, H. (2005). Teias alimentares: da conectividade à energia . (H. Caswell, Ed.). Avanços na pesquisa ecológica (Vol. 36). Elsevier Ltd. pp. 209
- Curtis, H. et ai. (2008). Biologia 7ª Edição. Buenos Aires-Argentina: Editorial Panamericana Médica. pp. 1160
- Kitching, RL (2000). Teias alimentares e habitats de contêineres: a história natural e a ecologia dos fitotelmata . Cambridge University Press. pp. 447
- Lindemann, RL (1942). O aspecto trófico – dinâmico da ecologia. Ecology , 23, 399-418.
- Pascual, M. e Dunne, JA (2006). Redes Ecológicas: Vinculando Estrutura à Dinâmica em Redes Alimentares. (M. Pascual e JA Dunne, Eds.) Estudos do Instituto Santa Fe nas Ciências da Complexidade . Oxford University Press. pp. 405