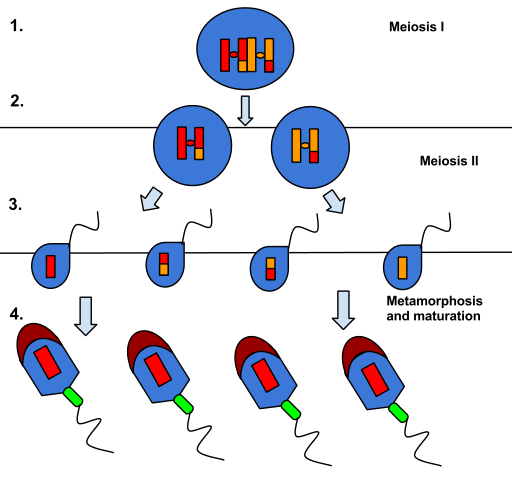
A espermatogênese é o processo de produção de espermatozoides nos testículos dos homens. Esse processo é fundamental para a reprodução sexual e envolve uma série de estágios distintos que ocorrem nas células germinativas masculinas. Durante a espermatogênese, as células germinativas passam por diversas divisões celulares e transformações morfológicas até se tornarem espermatozoides maduros, prontos para fertilizar um óvulo feminino. Neste processo, são formados espermatozoides haploides, com metade do material genético de uma célula somática normal. A espermatogênese é um processo complexo e altamente regulado que garante a produção contínua de espermatozoides ao longo da vida reprodutiva masculina.
Quais as principais características do processo de gametogênese nos organismos?
A gametogênese é o processo de formação dos gametas, células sexuais responsáveis pela reprodução dos organismos. Nos seres humanos, a espermatogênese é o processo de formação dos espermatozoides, células sexuais masculinas. Esse processo ocorre nos testículos e envolve uma série de estágios bem definidos.
Primeiramente, as células germinativas passam por uma fase de multiplicação chamada espermatogônia, na qual se dividem por mitose. Em seguida, as espermatogônias se transformam em espermatócitos primários, que por sua vez sofrem meiose, resultando em espermatócitos secundários. Estes, por sua vez, passam por uma segunda divisão meiótica, originando as espermátides.
As espermátides são as células precursoras dos espermatozoides e passam por um processo de maturação conhecido como espermiogênese. Durante a espermiogênese, as espermátides sofrem transformações morfológicas e se diferenciam em espermatozoides funcionais, prontos para a fertilização do óvulo.
É importante ressaltar que a espermatogênese é um processo contínuo e acontece ao longo da vida reprodutiva do homem. Além disso, a produção de espermatozoides é influenciada por diversos fatores, como hormônios, condições ambientais e estilo de vida.
Em resumo, a espermatogênese é um processo complexo e crucial para a reprodução dos organismos. Através da formação dos espermatozoides, os machos garantem a perpetuação da espécie e a diversidade genética. Portanto, a compreensão dos estágios e características desse processo é fundamental para o entendimento da biologia reprodutiva.
Espermatogênese: estágios e suas características
O espermatogénese é um processo de formação de espermatozóides de células germinativas (espermatogônias). Ocorre em indivíduos do sexo masculino de organismos eucarióticos com reprodução sexual .
Para que esse processo seja realizado com eficiência, ele precisa de condições específicas, incluindo: uma divisão cromossômica correta com expressões gênicas precisas e um meio hormonal adequado, para produzir um alto número de células funcionais.
A transformação do esperma em gametas maduros ocorre durante a maturação sexual nos organismos. Esse processo é desencadeado devido ao acúmulo de certos hormônios da gonadotrofina hipofisária, como o HCG (gonadotrofina coriônica humana) que está envolvida na produção de testosterona.
O que é espermatogênese?
A espermatogênese consiste na formação de gametas masculinos: esperma .
A produção dessas células sexuais começa nos túbulos seminíferos, localizados nos testículos. Esses túbulos ocupam cerca de 85% do volume total das gônadas e neles estão as células germinativas imaturas ou espermatogônias que são continuamente divididas por mitose.
Algumas dessas espermatogônias param de se reproduzir e se tornam espermatócitos primários, que iniciam o processo de meiose para cada um produzir um par de espermatócitos secundários com sua carga cromossômica completa.
Estes últimos completam o segundo estágio da meiose, dando origem finalmente a quatro espermatídeos com metade da carga cromossômica (haploides).
Posteriormente, sofrem alterações morfológicas, gerando espermatozóides, os quais são direcionados para o epidídimo localizado no escroto próximo aos testículos. Nesse ducto, ocorre a maturação dos gametas prontos para transmitir os genes do indivíduo.
O processo de espermatogênese depende da regulação hormonal e genética. Esse processo é dependente da testosterona; portanto, células especializadas (células de Leydig) são encontradas na produção desse hormônio nos túbulos seminíferos.
Elementos genéticos envolvidos
Alguns genes importantes na espermatogênese são o gene SF-1, que atua na diferenciação das células de Leydig, e o gene SRY, que intercede na diferenciação das células de Sertoli e na formação dos cordões testiculares. Outros genes estão envolvidos na regulação desse processo: RBMY, DBY, USP9Y e DAZ.
Este último é encontrado no cromossomo Y. Ele atua na codificação das proteínas de ligação ao RNA e sua ausência está ligada à infertilidade em alguns indivíduos.
Etapas e suas características
As células germinativas primárias (gonócitos) formam-se no saco vitelino e movem-se para a crista genital, dividindo-se entre as células de Sertoli, formando túbulos seminíferos. Os gonócitos são encontrados no interior, de onde migram para a membrana basal, dando origem à espermatogonia.
A proliferação primária de células germinativas e a formação de espermatozóides ocorrem durante o desenvolvimento embrionário do indivíduo. Logo após o nascimento, o processo de divisão mitótica dessas células é interrompido.
O processo pelo qual o esperma maduro é produzido é dividido em três fases: espermatogônica, espermatocítica e espermogênica.
1. Fase espermatogônica
À medida que o período de maturidade sexual dos indivíduos se aproxima, um aumento nos níveis de testosterona ativa a proliferação espermatogonal. Essas células germinativas se dividem para gerar uma série de espermatogônias que se diferenciam para formar os espermatócitos primários.
Nos seres humanos, vários tipos morfológicos de espermatogonia são distinguidos:
Anúncio de espermatogonia: localizado próximo às células intersticiais do túbulo seminífero. Eles sofrem divisões mitóticas que geram um par do tipo Ad que, por sua vez, continuam a se dividir, ou um par do tipo Ap.
Ap espermatogonia: seguem o processo de diferenciação para gerar espermatozóides, dividindo-se consecutivamente por mitose.
Espermatogonia B. Produto da divisão mitótica da espermatogonia Ap. Eles têm um núcleo esferoidal e a peculiaridade de estarem conectados entre si por “pontes citoplasmáticas”.
Eles formam um tipo de sincício que persiste em estágios subsequentes, separando-se na diferenciação espermática, quando os espermatozóides são liberados no lúmen do túbulo seminífero.
A união citoplasmática entre essas células permite o desenvolvimento sincronizado de cada par de espermatogônias e cada uma delas obtém as informações genéticas completas necessárias para sua operação, pois mesmo após a meiose, essas células continuam a se desenvolver.
2. Fase espermatocítica
Nesta fase, a espermatogonia B foi dividida mitoticamente, formando espermatócitos I (primários) que duplicam seus cromossomos, de modo que cada célula carrega dois conjuntos de cromossomos, transportando o dobro da quantidade usual de informações genéticas.
Posteriormente, são realizadas divisões meióticas desses espermatócitos, de modo que o material genético neles sofre reduções até atingir o caráter haplóide.
Mitose I
Na primeira divisão meiótica, os cromossomos são condensados na prófase e, no caso dos humanos, resultam 44 autossomos e dois cromossomos (um X e um Y), cada um com um conjunto de cromátides.
Os cromossomos homólogos se acasalam enquanto se alinham na placa equatorial da metáfase . Esses arranjos são chamados de tétrades porque contêm dois pares de cromatídeos.
Os tétrades trocam material genético (cruzamento), reorganizando as cromátides em uma estrutura chamada complexo sinaptonêmico.
Nesse processo, a diversificação genética ocorre quando as informações são trocadas entre cromossomos homólogos herdados do pai e da mãe, garantindo que todas as células espermáticas produzidas a partir delas sejam diferentes.
No final do cruzamento, os cromossomos são separados, movendo-se para pólos opostos do fuso meiótico, “dissolvendo” a estrutura das tetrades, mantendo juntos as cromátides recombinantes de cada cromossomo.
Outra maneira de garantir a diversidade genética em relação aos pais é pela distribuição aleatória dos cromossomos derivados do pai e da mãe para os pólos do fuso. No final desta divisão meiótica, são produzidos espermatócitos II (secundários).
Meiose II
Os espermatócitos secundários iniciam o processo da segunda meiose imediatamente após sua formação, sem sintetizar novo DNA . Como resultado, cada espermatozóide tem metade da carga cromossômica e cada cromossomo possui um par de cromátides irmãs com DNA duplicado.
Na metáfase, os cromossomos são distribuídos e alinhados na placa equatorial, e os cromátides se separam migrando para lados opostos do eixo meiótico.
Após a recomposição das membranas nucleares, o esperma haplóide é obtido com metade dos cromossomos (23 em humanos), uma cromatídeo e uma cópia da informação genética (DNA).
3. Fase espermiogênica
A espermiogênese é a última fase do processo de espermatogênese e não ocorre divisões celulares, mas alterações morfológicas e metabólicas que permitem que a diferenciação celular amadureça o esperma haplóide.
As alterações celulares ocorrem enquanto o esperma está ligado à membrana plasmática das células de Sertoli e podem ser descritas em quatro fases:
Fase de Golgi
É o processo pelo qual o aparelho de Golgi dá origem ao acrossoma, pelo acúmulo de grânulos proacrossomais ou PAS (ácido periódico-reativo de Schiff) nos complexos de Golgi.
Esses grânulos fluem para uma vesícula acrossômica localizada próxima ao núcleo e sua posição determina a porção anterior do esperma.
Os centríolos se movem em direção à porção posterior do esperma, alinhando-se perpendicularmente à membrana plasmática e fabricando os dupletos que integrarão os microtúbulos do axonema na base do flagelo do esperma.
Cap Phase
A vesícula acrosômica cresce e se estende sobre a porção anterior do núcleo, formando a tampa acrossoma ou acrossomal. Nesta fase, o conteúdo nuclear é condensado e a parte do núcleo que permanece sob o acrossoma engrossa, perdendo seus poros.
Fase Acrossômica
O núcleo se estende de redondo a elíptico, e o flagelo é orientado de modo que sua extremidade anterior adere às células de Sertoli, apontando para a lâmina basal dos túbulos seminíferos, dentro dos quais o flagelo se estende em formação.
O citoplasma viaja na direção posterior da célula e os microtúbulos citoplasmáticos se acumulam em uma bainha cilíndrica (manchette) que vai da tampa acrossômica até a porção posterior do esperma.
Depois que o flagelo se desenvolve, os centríolos retornam ao núcleo, aderindo a uma ranhura na porção posterior do núcleo, da qual partem nove fibras espessas que atingem os microtúbulos do axonema; dessa maneira, o núcleo e o flagelo estão conectados. Essa estrutura é conhecida como região do pescoço.
As mitocôndrias se movem em direção à região posterior do pescoço, circundando as fibras espessas e são dispostas em uma bainha helicoidal apertada, formando a região intermediária da cauda do esperma. O citoplasma se move para cobrir o flagelo já formado, e a “manchette” se dissolve.
Fase de maturação
O excesso de citoplasma é fagocitado pelas células de Sertoli, formando o corpo residual. A ponte citoplasmática que se forma na espermatogonia B permanece nos corpos residuais, de modo que os espermatozóides são separados.
Finalmente, os espermatozóides são liberados das células de Sertoli, liberando-se à luz do túbulo seminífero de onde são transportados através dos tubos retos, testículos e canais eferentes para o epidídimo.
Regulação hormonal
A espermatogênese é um processo finamente regulado por hormônios, principalmente a testosterona. Nos seres humanos, todo o processo é desencadeado na maturação sexual, pela liberação no hipotálamo do hormônio GnRH, que ativa a produção e o acúmulo de gonodotropinas hipofisárias (LH, FSH e HCG).
As células de Sertoli sintetizam proteínas transportadoras de testosterona (ABP) por estimulação do FSH e, juntamente com a testosterona liberada pelas células de Leydig (estimuladas pelo LH), garantem uma alta concentração desse hormônio nos túbulos seminíferos.
Nas células de Sertoli, o estradiol também é sintetizado, o que está envolvido na regulação da atividade das células de Leydig.
Fertilização
O epidídimo é conectado ao ducto deferente que flui para a uretra, permitindo finalmente que o esperma saia e, posteriormente, procurando um óvulo para fertilizar, completando o ciclo de reprodução sexual.
Uma vez liberado, o esperma pode morrer em questão de minutos ou horas, tendo que encontrar gameta feminino antes que isso aconteça.
Nos seres humanos, cerca de 300 milhões de espermatozóides são liberados em cada ejaculação durante a relação sexual, mas apenas cerca de 200 sobrevivem até atingirem a região onde podem acasalar.
O esperma deve passar por um processo de treinamento no trato reprodutivo feminino, onde adquire maior mobilidade do flagelo e prepara a célula para a reação do acrossoma. Essas características são necessárias para fertilizar os óvulos.
Treinamento de esperma
Entre as alterações que os espermatozóides apresentam, destacam-se modificações bioquímicas e funcionais, como hiperpolarização da membrana plasmática, aumento do pH citosólico, alterações nos lipídios e proteínas e ativação dos receptores da membrana, permitindo que sejam reconhecidos pela zona pelúcida Para participar deste.
Essa região funciona como uma barreira química para evitar o cruzamento de espécies, uma vez que, ao não reconhecer receptores específicos, a fertilização não é realizada.
Os óvulos têm uma camada de células granulares e são cercados por altas concentrações de ácido hialurônico que formam uma matriz extracelular. Para penetrar nesta camada de células, os espermatozóides têm enzimas hialuronidases.
Ao entrar em contato com a zona pelúcida, é desencadeada a reação do acrossoma, na qual o conteúdo da tampa acrossomal (como enzimas hidrolíticas) é liberado, o que ajuda o espermatozóide a atravessar a região e a se unir à membrana plasmática do óvulo, liberando dentro dele, seu conteúdo citoplasmático, organelas e núcleo.
Reação cortical
Em alguns organismos, ocorre uma despolarização da membrana plasmática do óvulo quando entra em contato com um espermatozóide, impedindo que mais de um fertilize.
Outro mecanismo para prevenir a polisspermia é a reação cortical, onde são liberadas enzimas que alteram a estrutura da zona pelúcida, inibindo a glicoproteína ZP3 e ativando a ZP2, tornando essa região impenetrável para outros espermatozóides.
Características do esperma
Os gametas masculinos possuem características que os tornam muito diferentes dos femininos e altamente adaptados para propagar os genes do indivíduo para as gerações seguintes.
Ao contrário dos óvulos, os espermatozóides são as menores células presentes no corpo e têm um flagelo que lhes permite mover-se para alcançar o gameta feminino (que não tem essa mobilidade) para fertilizá-lo. Este flagelo consiste em pescoço, região intermediária, região principal e região terminal.
No pescoço estão os centríolos e na região intermediária estão localizadas as mitocôndrias, responsáveis por fornecer a energia necessária para sua mobilidade.
Em geral, a produção de espermatozóides é muito alta, sendo muito competitivos entre si, pois apenas 25% fertilizam um gameta feminino.
Diferenças entre espermatogênese e ovogênese
A espermatogênese possui características que a diferenciam da oogênese:
-As células continuamente meiose a partir da maturação sexual do indivíduo, cada célula produzindo quatro gametas maduros em vez de um.
-O esperma amadurece após um processo complexo que começa após a meiose.
-Para a produção de um espermatozóide, ocorrem duas vezes mais divisões celulares do que na formação de um óvulo.
Referências
- Alberts, B., Johnson, A., Lewis, J., Raff, M., Roberth, K. e Walter, P. (2008). Biologia Molecular da Célula. Garland Science, Grupo Taylor e Francis.
- Creighton, TE (1999). Enciclopédia de biologia molecular. John Wiley e filhos, Inc.
- Hill, RW, Wyse, GA, & Anderson, M. (2012). Fisiologia Animal . Editores da Sinauer Associates, Inc..
- Kliman, RM (2016). Enciclopédia de Biologia Evolutiva . Imprensa acadêmica
- Marina, S. (2003) Avanços no conhecimento da espermatogênese, implicações clínicas. Revista Ibero-americana de Fertilidade. 20 (4), 213-225.
- Ross, MH, Pawlina, W. (2006). Histologia Editorial médico pan-americano.