As histonas são proteínas essenciais para a organização do DNA dentro do núcleo das células eucarióticas. Elas possuem uma estrutura globular e altamente básica, o que lhes permite interagir fortemente com os ácidos nucleicos. Existem cinco tipos principais de histonas: H1, H2A, H2B, H3 e H4. Cada tipo de histona possui funções específicas na compactação do DNA, regulação da expressão gênica e manutenção da estabilidade do genoma. Em conjunto, as histonas formam um complexo chamado de nucleossomo, que é a unidade básica da cromatina. As histonas desempenham um papel fundamental na regulação da expressão gênica e na organização tridimensional do genoma, sendo essenciais para a manutenção da integridade genômica e para o funcionamento adequado das células.
Conheça os tipos e funções das histonas no DNA eucarionte em detalhes.
As histonas são proteínas altamente conservadas que desempenham um papel fundamental na organização e compactação do DNA nos núcleos das células eucarióticas. Elas são compostas por um alto teor de aminoácidos básicos, como a lisina e a arginina, o que lhes confere uma carga positiva que interage fortemente com o DNA, que é negativamente carregado.
Existem cinco principais tipos de histonas: H1, H2A, H2B, H3 e H4. Estas histonas formam octâmeros chamados de nucleossomos, nos quais o DNA se enrola ao redor delas. O H1 atua como uma espécie de “fecho” do nucleossomo, ajudando na compactação do DNA.
A estrutura das histonas é altamente conservada ao longo da evolução, o que sugere a importância dessas proteínas para a organização do DNA. Elas são ricas em hélices alfa, que formam os domínios onde o DNA se liga.
As principais funções das histonas no DNA eucariótico incluem a compactação do DNA, a regulação da expressão gênica e a proteção do DNA contra danos. Além disso, as modificações pós-traducionais das histonas, como a acetilação e a metilação, podem afetar a estrutura do cromatina e, consequentemente, a atividade dos genes.
Em resumo, as histonas desempenham um papel crucial na organização e regulação do DNA nos núcleos das células eucarióticas, garantindo a expressão correta dos genes e a integridade do genoma.
Conheça os 4 tipos de cromossomos presentes nos seres humanos e suas funções.
As histonas são proteínas essenciais para a organização do DNA nos cromossomos. Elas possuem uma estrutura altamente conservada e são responsáveis por compactar e regular a expressão dos genes. Existem cinco tipos principais de histonas: H1, H2A, H2B, H3 e H4.
A histona H1 é responsável por estabilizar o DNA nos nucleossomos, enquanto as histonas H2A, H2B, H3 e H4 formam o núcleo do nucleossomo. Juntas, essas histonas ajudam a compactar o DNA em estruturas mais condensadas, facilitando a divisão celular e a transcrição dos genes.
Em relação aos cromossomos humanos, existem quatro tipos principais: os cromossomos autossômicos, os cromossomos sexuais X e Y, os cromossomos mitocondriais e os cromossomos nucleares. Os cromossomos autossômicos contêm a maior parte do material genético, enquanto os cromossomos sexuais determinam o sexo do indivíduo.
Os cromossomos mitocondriais são responsáveis pela produção de energia na célula, enquanto os cromossomos nucleares contêm a maior parte dos genes responsáveis pelas características hereditárias. Cada tipo de cromossomo desempenha funções específicas no organismo humano, garantindo a estabilidade genética e o funcionamento adequado das células.
O papel crucial das histonas na divisão celular: importância e função durante o processo.
As histonas são proteínas altamente básicas que desempenham um papel crucial na organização do DNA nas células. Elas são responsáveis por compactar e condensar o DNA, permitindo que ele caiba dentro do núcleo celular. Além disso, as histonas desempenham um papel fundamental na regulação da expressão gênica, controlando quais genes são ativados ou desativados em determinado momento.
Durante a divisão celular, as histonas desempenham uma função essencial na segregação do material genético. Elas ajudam a garantir que o DNA seja distribuído de forma precisa e equitativa entre as células filhas. Sem a presença adequada de histonas, a divisão celular pode ser comprometida, levando a erros na distribuição do DNA e possíveis consequências para a célula e o organismo como um todo.
Existem cinco principais tipos de histonas: H1, H2A, H2B, H3 e H4. Cada tipo de histona desempenha funções específicas na organização do DNA e na regulação da expressão gênica. Por exemplo, a histona H1 está envolvida na estabilidade da estrutura de cromatina, enquanto as histonas H2A, H2B, H3 e H4 estão envolvidas na compactação do DNA e na formação do nucleossomo, a unidade básica da cromatina.
Em resumo, as histonas desempenham um papel fundamental na divisão celular, garantindo a correta segregação do material genético e a regulação da expressão gênica. Sem a presença adequada de histonas, a célula não seria capaz de realizar sua divisão de forma eficiente e correta, o que poderia ter sérias consequências para o organismo como um todo.
Local de produção das histonas: onde ocorre a síntese dessas proteínas nucleares essenciais.
As histonas são proteínas nucleares essenciais para a organização do DNA no núcleo das células eucarióticas. Elas estão envolvidas no processo de compactação do DNA, formando a cromatina, e desempenham um papel fundamental na regulação da expressão gênica.
A síntese das histonas ocorre no núcleo das células, mais especificamente no citoplasma, onde os ribossomos livres são responsáveis pela tradução do RNA mensageiro que contém as informações genéticas para a produção dessas proteínas. Após a síntese, as histonas são transportadas para o núcleo, onde se ligam ao DNA e participam da formação da cromatina.
É importante ressaltar que as histonas são altamente conservadas ao longo da evolução, o que demonstra a sua importância para os processos celulares. Existem cinco tipos principais de histonas: H1, H2A, H2B, H3 e H4, cada uma com funções específicas na compactação do DNA e na regulação da expressão gênica.
Em resumo, as histonas são proteínas nucleares essenciais para a organização do DNA no núcleo das células eucarióticas. A síntese dessas proteínas ocorre no citoplasma, com a posterior migração para o núcleo, onde desempenham suas funções vitais para a integridade do genoma.
Histonas: características, estrutura, tipos e funções
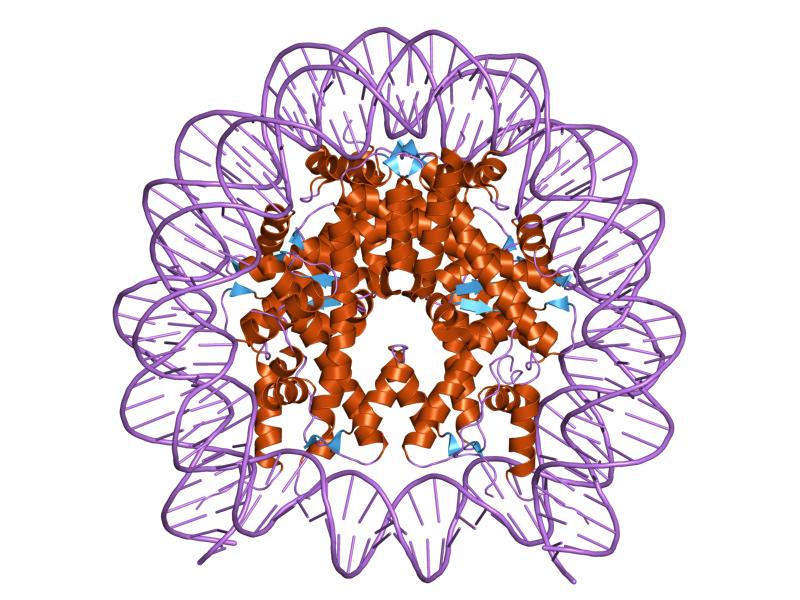
As histonas são proteínas básicas que interagem com o ADN para a formação de nucleossomas , que formam os fios constituintes da cromatina dos cromossomas em eucariotas .
Os nucleossomos, complexos formados por DNA e proteínas, foram descobertos em 1974 e são as histonas que montam esse nível basal de organização da cromatina. No entanto, a existência de proteínas histonas é conhecida desde antes da década de 1960.
As histonas são organizadas de tal maneira que o DNA de banda dupla é envolvido em torno de um centro de proteínas composto por essas proteínas que interagem estreitamente entre si. O centro de histonas tem a forma de um disco e o DNA leva cerca de 1,7 voltas ao seu redor.
Várias ligações de hidrogênio permitem que o DNA se ligue ao centro de proteínas formado por histonas em cada nucleossomo. Essas ligações são formadas, na maioria das vezes, entre os esqueletos de aminoácidos das histonas e o esqueleto de açúcar e fosfato do DNA. Algumas interações hidrofóbicas e ligações iônicas também participam .
As proteínas conhecidas como “complexos de remodelação da cromatina” são responsáveis pela quebra e formação das ligações de ligação entre o DNA e as histonas, permitindo a entrada de máquinas de transcrição no DNA contido nos nucleossomos.
Apesar da proximidade dos ácidos nucléicos ao centro proteico formado pelas histonas, eles estão dispostos de tal maneira que, se necessário, permitem a entrada de fatores de transcrição e outras proteínas relacionadas à expressão ou silenciamento de genes. .
As histonas podem sofrer várias modificações que geram múltiplas variantes, possibilitando a existência de muitas formas diferentes de cromatina que têm a propriedade de modular a expressão genética de diferentes maneiras.
Caracteristicas
São as proteínas eucarióticas mais preservadas da natureza. Foi demonstrado, por exemplo, que a histona de ervilha H4 difere em apenas duas das 102 posições de aminoácidos da proteína H4 de vaca.
Histonas são proteínas relativamente pequenas, com não mais que 140 aminoácidos. Eles são ricos em resíduos de aminoácidos básicos, portanto, possuem uma carga líquida positiva, o que contribui para sua interação com o ácido nucleico, carregado negativamente, para formar nucleossomos.
As histonas nucleossomais e de junção ou ponte são conhecidas. As histonas nucleossômicas são H3, H4, H2A e H2B, enquanto as histonas de ligação pertencem à família das histonas H1.
Durante a montagem do nucleossomo, os dímeros específicos H3-H4 e H2A-H2B são formados inicialmente. Dois dímeros H3-H4 são então unidos para formar tetrâmeros que posteriormente se combinam com os dímeros H2A-H2B, formando o centro octamérico.
Todas as histonas são sintetizadas principalmente durante a fase S do ciclo celular, e os nucleossomos são reunidos em hélices de DNA nascentes, logo após o garfo de replicação.
Estrutura
A estrutura geral das histonas inclui uma região básica de aminoácidos e uma região carboxila globular altamente conservada entre organismos eucarióticos.
Um motivo estrutural conhecido como “dobra de histonas”, composto por três hélices alfa conectadas por dois grampos de cabelo e formando um pequeno centro hidrofóbico, é responsável pelas interações proteína-proteína entre as histonas que formam o nucleossomo.
É essa dobra de histonas que forma o domínio carboxil globular das referidas proteínas nucleossômicas em todos os eucariotos.
As histonas também possuem pequenas “caudas” ou regiões amino-terminais e outras regiões carboxil-terminais (acessíveis a proteases), com no máximo 40 aminoácidos. Ambas as regiões são ricas em aminoácidos básicos que podem sofrer várias modificações covalentes pós-traducionais.
Histonas de junção
Nos eucariotos, existem duas famílias de histonas de ligação, diferenciadas uma da outra por sua estrutura. Alguns têm uma estrutura tripartida, com o domínio globular descrito acima flanqueado por domínios N-e C-terminais “não estruturados”; enquanto outros têm apenas um domínio C-terminal.
Embora a maioria das histonas seja conservada, durante a embriogênese específica ou a maturação de células especializadas em alguns organismos, algumas variantes específicas podem surgir. Algumas variações estruturais estão relacionadas a modificações pós-traducionais, como as seguintes:
– Fosforilação : acredita-se estar relacionada à modificação do grau de condensação da cromatina e é comumente encontrada em resíduos de serina.
– Acetilação : associada a regiões cromossômicas que são transcricionalmente ativas. Ocorre normalmente nas cadeias laterais dos resíduos de lisina. Quando ocorrem nesses resíduos, sua carga positiva diminui, diminuindo a afinidade das proteínas pelo DNA.
– Metilação : pode ocorrer como mono-, di ou trimetilação dos resíduos de lisina que se projetam do núcleo da proteína.
Enzimas específicas são responsáveis por fazer essas modificações covalentes nas histonas. Essas enzimas incluem histona acetil transferases (HATs), complexos de histona desacetilase (HDACs) e histona metiltransferases e desmetilases.
Tipos
A caracterização das histonas foi realizada por várias técnicas bioquímicas, dentre as quais destacam-se cromatografias baseadas em resinas de troca catiônica fraca.
Certos autores estabelecem uma forma de classificação na qual 5 tipos principais de histonas são diferenciados em eucariotos: FI, com proteínas de 21 kDa; F2A1 ou FIV, com mais ou menos 11,3 kDa; F2A2 ou FIIbI, 14,5 kDa; F2B ou FIIb2, com um peso molecular de 13,7 kDa, e F3 ou FIII, de 15,3 kDa.
Todos esses tipos de histonas, com exceção do grupo IF, são encontrados em quantidades equimolares nas células.
Outra classificação, com a mesma validade e talvez a mais usada atualmente, propõe a existência de dois tipos diferentes de histonas, a saber: aquelas que fazem parte do octâmero do nucleossomo e as histonas de junção ou ponte que ligam os nucleossomos entre sim
Algumas variantes também podem ocorrer entre as espécies e, diferentemente das histonas centrais, as variantes são sintetizadas durante a interface e inseridas na cromatina pré-formada por meio de um processo dependente da energia liberada pela hidrólise do ATP.
Histonas nucleossomais
O centro de um nucleossomo consiste em um par de cada uma das quatro histonas constituintes: H2a, H2b, H3 e H4; em que segmentos de DNA de cerca de 145 pares de bases são rolados.
As histonas H4 e H2B são, em princípio, invariáveis. Algumas variações são evidentes, no entanto, nas histonas H3 e H2A, cujas propriedades biofísicas e bioquímicas alteram a natureza normal do nucleossomo.
Variante da histona H2A em humanos, a proteína H2A.Z possui uma grande região ácida e pode favorecer a estabilidade do nucleossomo, dependendo das variantes da histona H3 às quais está associada.
Essas histonas mostram alguma variabilidade entre as espécies, com um caso especial sendo o da histona H2B, para a qual o primeiro terço da molécula é altamente variável.
Histonas de junção
Histonas de ligação ou ponte são histonas da classe H1. Eles são responsáveis pela união entre nucleossomos e pela proteção do DNA que se destaca no início e no final de cada partícula.
Diferentemente das histonas nucleossômicas, nem todas as histonas do tipo H1 possuem a região globular da “dobra” das histonas. Essas proteínas se ligam ao DNA entre os nucleossomos, facilitando uma mudança no equilíbrio da cromatina em direção a um estado mais condensado e menos ativo, transcricionalmente.
Os estudos associaram essas histonas ao envelhecimento, reparo do DNA e processos apoptóticos, portanto, acredita-se que eles tenham um papel crucial na manutenção da integridade genômica.
Funções
Todos os resíduos de aminoácidos das histonas participam, de uma maneira ou de outra, de sua interação com o DNA, o que explica o fato de serem tão conservados entre os reinos dos organismos eucarióticos.
A participação de histonas na embalagem de DNA em forma de cromatina é de grande relevância para organismos multicelulares complexos nos quais diferentes linhagens celulares podem se especializar apenas alterando a acessibilidade de seus genes à maquinaria transcricional.
As regiões genômicas transcricionalmente ativas são densas nos nucleossomos, sugerindo que a associação do DNA às proteínas histonas é crucial para a regulação negativa ou positiva de sua transcrição.
Da mesma forma, ao longo da vida de uma célula, a resposta a um grande número de estímulos, internos e externos, depende de pequenas alterações na cromatina, que normalmente têm a ver com remodelação e modificação pós-traducional das histonas encontradas em estreita relação com o DNA.
Múltiplas variáveis histonas exercem funções diferentes nos eucariotos. Um deles tem a ver com a participação de uma variante da histona H3 na formação das estruturas centroméricas responsáveis pela segregação dos cromossomos durante a mitose.
Foi demonstrado que a contraparte desta proteína em outros eucariotos é essencial para a montagem da proteína cinetocora à qual os microtúbulos do fuso se ligam durante a mitose e meiose .
Referências
- Alberts, B., Johnson, A., Lewis, J., Morgan, D., Raff, M., Roberts, K. e Walter, P. (2015). Biologia Molecular da Célula (6a ed.). Nova York: Garland Science.
- Campos, EI, & Reinberg, D. (2009). Histons: anotando cromatina. Annu Rev. Genet. , 43 , 559-599.
- Harvey, AC, & Downs, JA (2004). Que funções as histonas do vinculador fornecem? Molecular Microbiology , 53 , 771-775.
- Henikoff, S., & Ahmad, K. (2005). Montagem de histonas variantes na cromatina. Annu Rev. Cell. Dev. Biol. , 21 , 133-153.
- Isenberg, I. (1979). Histons Annu Rev. Biochem. , 48 , 159-191.
- Kornberg, RD, e Thomas, JO (1974). Estrutura da cromatina: oligômeros das histonas. Science , 184 (4139), 865-868.
- Smith, E., DeLange, R. & Bonner, J. (1970). Química e Biologia das Histonas. Revisões fisiológicas , 50 (2), 159-170.