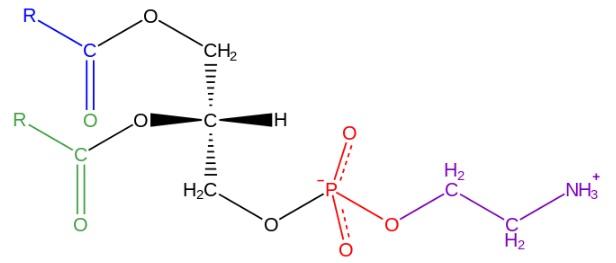
A fosfatidiletanolamina é um fosfolipídeo essencial encontrado nas membranas celulares de todos os organismos vivos. Sua estrutura consiste em um grupo fosfato, dois ácidos graxos e um grupo etanolamina ligados a uma molécula de glicerol. A biossíntese da fosfatidiletanolamina ocorre principalmente na via da CDP-etanolamina, onde a etanolamina é adicionada ao fosfatidilcolina. A fosfatidiletanolamina desempenha importantes funções no organismo, como a manutenção da integridade das membranas celulares, sinalização celular e regulação do metabolismo lipídico.
Qual a importância da fosfatidiletanolamina no organismo?
A fosfatidiletanolamina é um fosfolipídio essencial para o funcionamento adequado do organismo. Presente em todas as membranas celulares, ela desempenha diversas funções vitais para a saúde do corpo.
Uma das principais funções da fosfatidiletanolamina é garantir a integridade e a fluidez das membranas celulares. Isso é fundamental para a comunicação entre as células, o transporte de nutrientes e a proteção contra substâncias nocivas.
Além disso, a fosfatidiletanolamina também está envolvida na regulação do metabolismo lipídico e na sinalização celular. Ela é essencial para a produção de lipoproteínas, que são responsáveis pelo transporte de gorduras no sangue.
Outra função importante da fosfatidiletanolamina é a sua participação na coagulação sanguínea. Ela atua como um cofator em diversas reações enzimáticas que levam à formação de coágulos, garantindo a hemostasia e evitando hemorragias.
Em resumo, a fosfatidiletanolamina é um componente fundamental para a estrutura e o funcionamento das células do organismo. Sua presença é indispensável para a manutenção da saúde e do equilíbrio do corpo.
Local de produção dos fosfolipídios no organismo humano: você sabe onde é?
A fosfatidiletanolamina é um tipo de fosfolipídio presente em membranas celulares e desempenha diversas funções fisiológicas no organismo humano. Mas você sabe onde esses compostos são produzidos?
Os fosfolipídios, incluindo a fosfatidiletanolamina, são sintetizados principalmente no retículo endoplasmático das células. Nesse organelo, ocorrem diversas reações bioquímicas que resultam na formação desses importantes componentes das membranas celulares.
A biossíntese da fosfatidiletanolamina envolve várias etapas, incluindo a adição de grupos fosfato e etanolamina a um esqueleto de glicerol. Essas reações são catalisadas por enzimas específicas presentes no retículo endoplasmático.
Além disso, a fosfatidiletanolamina desempenha um papel fundamental na estrutura e função das membranas celulares, atuando na regulação da permeabilidade e na sinalização celular. Sua presença é essencial para a integridade e funcionamento adequado das células do organismo humano.
Portanto, conhecer o local de produção dos fosfolipídios, como a fosfatidiletanolamina, é fundamental para compreender a importância desses compostos para a saúde e o funcionamento do organismo. A biossíntese dessas moléculas no retículo endoplasmático é um processo complexo e essencial para a manutenção da homeostase celular.
Fosfatidiletanolamina: estrutura, biossíntese e funções
A fosfatidiletanolamina (PE) é um abundande glicerofosfolípido nas membranas plasmáticas de procariotas . Pelo contrário, nas membranas celulares eucarióticas , este é o segundo glicerofosfolípido mais abundante na face interna da membrana plasmática após a fosfatidilcolina.
Não obstante a abundância de fosfatidiletanolamina, sua abundância depende não apenas do tipo de célula, mas também do compartimento e do tempo do ciclo de vida celular específico considerado.
Membranas biológicas são barreiras que definem organismos celulares. Eles não apenas possuem funções de proteção e isolamento, mas também são essenciais para o estabelecimento de proteínas que requerem um ambiente hidrofóbico para o funcionamento ideal.
Tanto os eucariotos quanto os procariontes possuem membranas compostas principalmente por glicerofosfolípides e, em menor grau, esfingolipídios e esteróis.
Glicerofosfolípides são moléculas anfipáticas estruturadas em um esqueleto de L-glicerol que é esterificado nas posições sn- 1 e sn- 2 por dois ácidos graxos de comprimento e grau de saturação variados. No hidroxil da posição sn- 3 , é esterificado por um grupo fosfato, ao qual, por sua vez, diferentes tipos de moléculas que dão origem às várias classes de glicerofosfolípides podem ser ligados.
No mundo celular, existe uma grande variedade de glicerofosfolipídios, no entanto, os mais abundantes são fosfatidilcolina (PC), fosfatidiletanolamina (PE), fosfatidilserina (PS), fosfatidilinositol (PI), ácido fosfatídico (PA), fosfatidilglicerol (PG) e cardiolipina (CL).
Estrutura
A estrutura da fosfatidiletanolamina foi descoberta por Baer et al em 1952. Como foi determinado experimentalmente para todos os glicerofosfolípides, a fosfatidiletanolamina é formada por uma molécula de glicerol esterificado nas posições sn -1 e sn -2 com cadeias ácidas ácidos graxos entre 16 e 20 átomos de carbono.
Os ácidos graxos esterificados no hidroxil sn- 1 são geralmente saturados (sem ligações duplas) com comprimentos máximos de 18 átomos de carbono, enquanto as cadeias ligadas na posição sn- 2 são mais longas e com uma ou mais insaturações ( links duplos).
O grau de saturação dessas cadeias contribui para a elasticidade da membrana, que exerce grande influência na inserção e seqüestro de proteínas na bicamada.
A fosfatidiletanolamina é considerada um glicerofosfolípido não lamelar, por apresentar uma forma geométrica cônica. Essa forma é dada pelo tamanho pequeno do seu grupo polar ou “cabeça”, em relação ao das cadeias de ácidos graxos que compreendem as “caudas” hidrofóbicas.
A “cabeça” ou grupo polar da fosfatidiletanolamina tem um caráter zwitteriônico, ou seja, possui grupos que podem ser carregados positiva e negativamente sob certas condições de pH.
Esse recurso permite estabelecer ligações de hidrogênio com uma grande quantidade de resíduos de aminoácidos e sua distribuição de carga é um determinante essencial para a topologia dos domínios de muitas proteínas integrais da membrana.
Biossíntese
Nas células eucarióticas, a síntese de lipídios estruturais é geograficamente restrita, sendo o principal local da biossíntese o retículo endoplasmático (ER) e, em menor grau, o aparelho de Golgi .
Existem quatro rotas biossintéticas independentes para a produção de fosfatidiletanolamina: (1) a rota CDP-etanolamina, também conhecida como rota Kennedy; (2) a via PSD para descarboxilação da fosfatidilserina (PS); (3) a acilação da liso-PE e (4) as reações de mudança de base do grupo polar de outros glicerofosfolípides.
Rota Kennedy
A biossíntese de fosfatidiletanolamina por esta via é limitada a ER e foi demonstrado que nas células hepáticas de Hamster é a principal via de produção. Consiste em três etapas enzimáticas consecutivas catalisadas por três enzimas diferentes.
Na primeira etapa, a fosfoetanolamina e o ADP são produzidos graças à ação da etanolamina cinase, que catalisa a fosforilação da etanolamina dependente de ATP .
Diferentemente das plantas, nem mamíferos nem leveduras são capazes de produzir esse substrato; portanto, ele deve ser consumido na dieta ou obtido pela degradação de moléculas de fosfatidiletanolamina ou esfingosina pré-existentes.
A fosfoetanolamina é usada pela CTP: fosfoetanolamina citidiltransferase (ET) para formar o composto de alta energia CDP: etanolamina e um fosfato inorgânico.
A 1,2-diacilglicerol etanolamina fosfotransferase (ETP) usa a energia contida na ligação CDP-etanolamina para ligar covalentemente a etanolamina a uma molécula de diacilglicerol inserida na membrana, resultando em fosfatidiletanolamina.
Caminho PSD
Esta rota opera tanto em procariontes quanto em leveduras e mamíferos. Nas bactérias, ocorre na membrana plasmática, mas nos eucariotos, ocorre em uma área do retículo endoplasmático que possui estreita relação com a membrana mitocondrial.
Nos mamíferos, a via é catalisada por uma única enzima, a fosfatidilserina descarboxilase (PSD1p), que é incorporada na membrana mitocondrial, cujo gene é codificado pelo núcleo. A reação envolve descarboxilação de PS em fosfatidiletanolamina.
As duas vias restantes (acilação do liso-PE e troca do grupo polar dependente de cálcio) ocorrem no retículo endoplasmático, mas não contribuem significativamente para a produção total de fosfatidiletanolamina em células eucarióticas.
Funções
Os glicerofosfolípides têm três funções principais na célula, dentre as quais se destacam as funções estruturais, de armazenamento de energia e de sinalização celular.
A fosfatidiletanolamina está associada à ancoragem, estabilização e dobra de múltiplas proteínas de membrana, bem como às alterações conformacionais necessárias ao funcionamento de muitas enzimas.
Existem evidências experimentais que propõem a fosfatidiletanolamina como um glicerofosfolípido crucial na fase tardia da telófase, durante a formação do anel contrátil e o estabelecimento do fragmoplasto que permite a divisão da membrana das duas células filhas.
Também desempenha um papel importante em todos os processos de fusão e fissão (união e separação) das membranas do retículo endoplasmático e do aparelho de Golgi.
Em E. coli , foi comprovado que a fosfatidiletanolamina é necessária para o correto dobramento e função da enzima permease lactose, por isso foi sugerido que ele desempenha um papel de “acompanhante” molecular.
A fosfatidiletanolamina é o principal doador da molécula de etanolamina necessária para a modificação pós-traducional de inúmeras proteínas, como as âncoras GPI.
Este glicerofosfolípido é um precursor de numerosas moléculas com atividade enzimática. Além disso, moléculas derivadas de seu metabolismo, bem como diacilglicerol, ácido fosfatídico e alguns ácidos graxos, podem atuar como segundos mensageiros. Além disso, é um substrato importante para a produção de fosfatidilcolina.
Referências
- Brouwers, JFHM, Vernooij, EAAM, Tielens, AGM e van Golde, LMG (1999). Rápida separação e identificação de espécies moleculares de fosfatidiletanolamina. Journal of Lipid Research, 40 (1), 164-169. Recuperado de jlr.org
- Calzada, E., McCaffery, JM, & Claypool, SM (2018). A fosfatidiletanolamina produzida na membrana mitocondrial interna é essencial para a função complexa do citocromo bc1 da levedura 3. BioRxiv, 1, 46.
- Calzada, E., Onguka, O. & Claypool, SM (2016). Metabolismo da fosfatidiletanolamina na saúde e na doença. Revista Internacional de Biologia Celular e Molecular (Vol. 321). Elsevier Inc.
- Gibellini, F., & Smith, TK (2010). A síntese de via via de novo de fosfatidiletanolamina e fosfatidilcolina. IUBMB Life, 62 (6), 414-428.
- Harayama, T. & Riezman, H. (2018). Compreendendo a diversidade da composição lipídica da membrana. Nature Reviews Molecular Cell Biology, 19 (5), 281-296.
- Luckey, M. (2008). Biologia estrutural de membranas: com fundamentos bioquímicos e biofísicos. Cambrudge University Press. Recuperado de cambrudge.org
- Seddon, JM, Cevc, G., Kaye, RD, e Marsh, D. (1984). Estudo de difração de raios X do polimorfismo de diacil e dialquilfosfatidiletanolaminas hidratadas. Bioquímica, 23 (12), 2634-2644.
- Sendecki, AM, Poyton, MF, Baxter, AJ, Yang, T. e Cremer, PS (2017). Bicamadas lipídicas suportadas com fosfatidiletanolamina como componente principal. Langmuir, 33 (46), 13423-13429.
- van Meer, G., Voelker, DR, & Feignenson, GW (2008). Lipídios da membrana: onde estão e como se comportam. Nature Reviews, 9, 112-124.
- Vance, JE (2003). Biologia Molecular e Celular do Metabolismo da Fosfatidilserina e Fosfatidiletanolamina. Em K. Moldave (Ed.), Progress Nucleic Acid Research and Molecular Biology (pp. 69-111). Imprensa acadêmica
- Vance, JE (2008). Fosfatidilserina e fosfatidiletanolamina em células de mamíferos: dois aminofosfolípidos metabolicamente relacionados. Journal of Lipid Research, 49 (7), 1377-1387.
- Vance, JE, & Tasseva, G. (2013). Formação e função de fosfatidilserina e fosfatidiletanolamina em células de mamíferos. Biochimica et Biophysica Acta – Biologia Molecular e Celular de Lipídios, 1831 (3), 543–554.
- Watkins, SM, Zhu, X. e Zeisel, SH (2003). A atividade da fosfatidiletanolamina-N-metiltransferase e a colina na dieta regulam o fluxo lipídico do plasma hepático e o metabolismo essencial dos ácidos graxos em camundongos. The Journal of Nutrition, 133 (11), 3386-3391.